Ahli medis artikel

Publikasi baru
Kolera vibrio
Terakhir ditinjau: 04.07.2025

Semua konten iLive ditinjau secara medis atau diperiksa fakta untuk memastikan akurasi faktual sebanyak mungkin.
Kami memiliki panduan sumber yang ketat dan hanya menautkan ke situs media terkemuka, lembaga penelitian akademik, dan, jika mungkin, studi yang ditinjau secara medis oleh rekan sejawat. Perhatikan bahwa angka dalam tanda kurung ([1], [2], dll.) Adalah tautan yang dapat diklik untuk studi ini.
Jika Anda merasa salah satu konten kami tidak akurat, ketinggalan zaman, atau dipertanyakan, pilih dan tekan Ctrl + Enter.
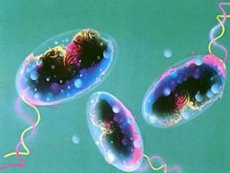
Menurut WHO, kolera adalah penyakit menular yang ditandai dengan diare akut, parah, dan dehidrasi dengan tinja berupa air beras, yang merupakan konsekuensi dari infeksi Vibrio cholerae. Karena penyakit ini ditandai dengan kemampuan yang jelas untuk menyebar secara luas dalam bentuk epidemi, perjalanan penyakit yang parah, dan tingkat kematian yang tinggi, kolera dianggap sebagai infeksi yang sangat berbahaya.
Tanah air kolera secara historis adalah India, atau lebih tepatnya, delta sungai Gangga dan Brahmaputra (sekarang India Timur dan Bangladesh), tempat kolera telah ada sejak dahulu kala (wabah kolera di wilayah ini diamati sejak 500 SM). Keberadaan fokus endemik kolera yang lama di sini dijelaskan oleh banyak alasan. Vibrio kolera tidak hanya dapat bertahan hidup di air untuk waktu yang lama, tetapi juga berkembang biak di dalamnya dalam kondisi yang menguntungkan - suhu di atas 12 °C, adanya bahan organik. Semua kondisi ini terbukti di India: iklim tropis (suhu tahunan rata-rata dari 25 hingga 29 °C), curah hujan dan rawa yang melimpah, kepadatan penduduk yang tinggi, terutama di delta Sungai Gangga, sejumlah besar bahan organik di dalam air, polusi air terus menerus sepanjang tahun dengan limbah dan kotoran, standar hidup material yang rendah dan ritual keagamaan dan pemujaan penduduk yang unik.
Dalam sejarah epidemi kolera, empat periode dapat dibedakan.
Periode I - hingga 1817, ketika kolera terkonsentrasi hanya di Asia Timur dan Selatan, terutama di India, dan tidak menyebar ke luar perbatasannya.
Periode II - dari tahun 1817 hingga 1926. Dengan terjalinnya hubungan ekonomi dan hubungan lainnya yang luas antara India dan negara-negara Eropa dan negara-negara lainnya, kolera menyebar ke luar India dan menyebar melalui jalur hubungan ekonomi dan agama, menyebabkan 6 pandemi yang merenggut jutaan nyawa manusia. Rusia adalah negara Eropa pertama yang dilanda kolera. Dari tahun 1823 hingga 1926, Rusia mengalami 57 tahun kolera. Selama kurun waktu tersebut, lebih dari 5,6 juta orang jatuh sakit karena kolera dan 2,14 juta orang meninggal karenanya ("40%).
Periode III - dari tahun 1926 hingga 1961 Kolera kembali menjadi fokus endemik utamanya, dan periode kesejahteraan relatif pun dimulai. Tampaknya dengan pengembangan sistem modern untuk membersihkan air minum, membuang dan mendisinfeksi air limbah, dan mengembangkan langkah-langkah anti-kolera khusus, termasuk pembentukan layanan karantina, negara-negara di dunia akan terlindungi secara andal dari invasi kolera lainnya.
Periode keempat dimulai pada tahun 1961 dan berlanjut hingga hari ini. Pandemi ketujuh dimulai bukan di India, tetapi di Indonesia, dengan cepat menyebar ke Filipina, Cina, negara-negara Indochina, dan kemudian ke negara-negara lain di Asia, Afrika, dan Eropa. Kekhasan pandemi ini meliputi fakta bahwa, pertama, itu disebabkan oleh varian khusus dari kolera vibrio - V. cholerae eltor, yang hingga tahun 1961 bahkan tidak secara resmi diakui sebagai agen penyebab kolera; kedua, dalam hal durasi, itu melampaui semua pandemi sebelumnya; ketiga, itu terjadi dalam dua gelombang, yang pertama berlangsung hingga tahun 1990, dan yang kedua dimulai pada tahun 1991 dan mencakup banyak negara di Amerika Selatan dan Utara, termasuk Amerika Serikat, yang belum pernah melihat epidemi kolera sejak tahun 1866. Dari tahun 1961 hingga 1996, 3.943.239 orang jatuh sakit karena kolera di 146 negara.
Agen penyebab kolera, Vibrio cholerae, ditemukan pada tahun 1883 selama pandemi kelima oleh R. Koch, tetapi vibrio pertama kali ditemukan dalam tinja pasien diare pada tahun 1854 oleh F. Pacini.
V. cholerae termasuk dalam famili Vibrionaceae, yang mencakup beberapa genus (Vibrio, Aeromonas, Plesiomonas, Photobacterium). Genus Vibrio memiliki lebih dari 25 spesies sejak tahun 1985, yang paling penting bagi manusia adalah V. cholerae, V. parahaemolyticus, V. alginolyticus, V. vulnificus, dan V. fluvialis.
Fitur utama dari genus Vibrio
Batang gram negatif pendek, tidak membentuk spora dan kapsul, melengkung atau lurus, berdiameter 0,5 µm dan panjang 1,5-3,0 µm, motil (V. cholerae bersifat monotrik, beberapa spesies memiliki dua atau lebih flagela polar); tumbuh dengan baik dan cepat pada media biasa, bersifat kemoorganotrof, dan memfermentasi karbohidrat untuk menghasilkan asam tanpa gas (glukosa difermentasi melalui jalur Embden-Meyerhof). Oksidase positif, membentuk indol, mereduksi nitrat menjadi nitrit (V. cholerae memberikan reaksi nitrosoindol positif), memecah gelatin, sering memberikan reaksi Voges-Proskauer positif (yaitu, membentuk asetilmetilkarbinol), tidak memiliki urease, tidak membentuk H2S, memiliki dekarboksilase lisin dan ornitin, tetapi tidak memiliki arginin dihidrolase. Ciri khas genus Vibrio adalah sensitivitas sebagian besar strain bakteri terhadap obat 0/129 (2,4-diamino-6,7-diazopropylpteridine), sementara perwakilan dari famili Pseudomonadaceae dan Enterobacteriaceae resisten terhadap obat ini. Vibrio bersifat aerob dan anaerob fakultatif, suhu optimum untuk pertumbuhan adalah 18-37 C, pH 8,6-9,0 (tumbuh dalam kisaran pH 6,0-9,6), beberapa spesies (halofil) tidak tumbuh tanpa adanya NaCl. Kandungan G + C dalam DNA adalah 40-50 mol % (untuk V. cholerae sekitar 47 mol %). Uji biokimia digunakan untuk membedakan dalam famili Vibrionaceae dari genus Aeromonas dan Plesiomonas yang secara morfologis mirip, serta untuk membedakan dari famili Enterobacteriaceae.
Vibrio cholera berbeda dari famili Pseudomonadaceae karena memfermentasi glukosa hanya melalui jalur Embden-Meyerhof (tanpa partisipasi O2), sedangkan yang pertama mengonsumsi glukosa hanya dengan adanya O2. Perbedaan di antara keduanya mudah terungkap pada media Hugh-Leifson. Media tersebut mengandung agar nutrisi, glukosa, dan indikator. Penaburan dilakukan dalam dua kolom dengan media Hugh-Leifson, yang salah satunya diisi dengan petroleum jelly (untuk menciptakan kondisi anaerobik). Dalam kasus pertumbuhan vibrio cholera, warna media berubah di kedua tabung reaksi, dalam kasus pertumbuhan pseudomonad - hanya dalam tabung reaksi tanpa petroleum jelly (kondisi pertumbuhan aerobik).
Vibrio cholera sangat tidak menuntut media nutrisi. Ia bereproduksi dengan baik dan cepat dalam air pepton (PV) alkali 1% (pH 8,6-9,0) yang mengandung 0,5-1,0% NaCl, melampaui pertumbuhan bakteri lain. Untuk menekan pertumbuhan Proteus, disarankan untuk menambahkan kalium tellurit (dalam pengenceran akhir 1:100.000) ke PV 1%. PV 1% adalah media pengayaan terbaik untuk vibrio cholera. Selama pertumbuhan, ia membentuk lapisan tipis, longgar, keabu-abuan pada permukaan PV setelah 6-8 jam, yang mudah hancur saat dikocok dan jatuh ke dasar dalam bentuk serpihan, PV menjadi agak keruh. Berbagai media selektif telah diusulkan untuk isolasi vibrio cholera: agar alkali, agar empedu-garam, albuminat alkali, agar alkali dengan darah, laktosa-sukrosa dan media lainnya. Yang terbaik adalah media TCBS (thiosulfate citrate-bromothymol sucrose agar) dan modifikasinya. Namun, MPA alkali paling sering digunakan, di mana vibrio kolera membentuk koloni berbentuk cakram yang halus, bening seperti kaca, berwarna kebiruan, dan memiliki konsistensi kental.
Bila disemai dengan cara disuntikkan ke dalam kolom gelatin, vibrio, setelah 2 hari pada suhu 22-23 C, menyebabkan pencairan dari permukaan dalam bentuk gelembung, kemudian berbentuk corong, dan akhirnya, lapis demi lapis.
Di dalam susu, vibrio berkembang biak dengan cepat, menyebabkan pembekuan setelah 24-48 jam, kemudian terjadi peptonisasi susu, dan setelah 3-4 hari vibrio mati karena terjadi pergeseran pH susu ke sisi asam.
B. Heiberg, berdasarkan kemampuannya memfermentasi manosa, sukrosa, dan arabinosa, membagi semua vibrio (kolera dan yang mirip kolera) ke dalam beberapa kelompok, yang jumlahnya sekarang berjumlah 8.
Vibrio cholerae termasuk kelompok pertama Heiberg.
Vibrio yang mirip dalam ciri morfologi, kultur, dan biokimia dengan vibrio kolera dulu dan sekarang disebut secara berbeda: paracholera, mirip kolera, vibrio NAG (vibrio non-aglutinasi); vibrio yang tidak termasuk dalam kelompok O1. Nama terakhir paling akurat menekankan hubungan mereka dengan vibrio kolera. Seperti yang ditetapkan oleh A. Gardner dan K. Venkat-Raman, vibrio kolera dan mirip kolera memiliki antigen H yang sama, tetapi berbeda dalam antigen O. Menurut antigen O, vibrio kolera dan mirip kolera saat ini dibagi menjadi 139 serogroup O, tetapi jumlahnya terus bertambah. Vibrio kolera termasuk dalam kelompok O1. Ia memiliki antigen A yang umum dan dua antigen spesifik tipe - B dan C, yang membedakan tiga serotipe V. cholerae - serotipe Ogawa (AB), serotipe Inaba (AC) dan serotipe Hikoshima (ABC). Vibrio cholerae dalam tahap disosiasi memiliki antigen OR. Dalam hal ini, serum O, serum OR dan serum spesifik tipe Inaba dan Ogawa digunakan untuk mengidentifikasi V. cholerae.
Pada tahun 1992-1993, epidemi kolera besar dimulai di Bangladesh, India, Cina, Malaysia, dan negara-negara lain, yang agen penyebabnya adalah serovar baru spesies Vibrio cholerae yang sebelumnya tidak dikenal. Ia berbeda dari V. cholerae O1 dengan fitur antigenik: ia memiliki antigen 0139 dan kapsul polisakarida dan tidak diaglutinasi oleh serum O lainnya. Semua sifat morfologi dan biologis lainnya, termasuk kemampuan untuk menyebabkan kolera, yaitu, untuk mensintesis kolerogen eksotoksin, ternyata mirip dengan sifat-sifat V. cholerae O1. Akibatnya, agen penyebab kolera baru, V. cholerae 0139, tampaknya muncul sebagai akibat dari mutasi yang mengubah antigen O. Ia diberi nama V. cholerae 0139 bengal.
Pertanyaan tentang hubungan antara apa yang disebut vibrio mirip kolera dengan V. cholerae telah lama tidak jelas. Namun, perbandingan antara V. cholerae dan vibrio mirip kolera (nag-vibrio) dengan lebih dari 70 fitur mengungkapkan kesamaan mereka sebesar 90%, dan tingkat homologi DNA antara V. cholerae dan vibrio nag-vibrio yang diteliti adalah 70-100%. Oleh karena itu, vibrio mirip kolera digabungkan menjadi satu spesies dengan vibrio kolera, yang membedakannya terutama pada antigen O, sehubungan dengan itu mereka disebut vibrio dari kelompok non-01 - V. cholerae non-01.
Spesies V. cholerae terbagi menjadi 4 biotipe: V. cholerae, V. eltor, V. proteus dan V. albensis. Sifat vibrio El Tor telah diperdebatkan selama bertahun-tahun. Vibrio ini diisolasi pada tahun 1906 oleh F. Gottschlich di stasiun karantina El Tor dari tubuh seorang peziarah yang meninggal karena disentri. F. Gottschlich mengisolasi beberapa strain tersebut. Mereka tidak berbeda dari vibrio kolera dalam semua sifat mereka dan diaglutinasi oleh serum O kolera. Namun, karena tidak ada kolera di antara para peziarah pada saat itu, dan pembawaan jangka panjang vibrio kolera dianggap tidak mungkin, pertanyaan tentang kemungkinan peran etiologi V. eltor dalam kolera tetap kontroversial untuk waktu yang lama. Selain itu, vibrio El Tor, tidak seperti V. cholerae, memiliki efek hemolitik. Akan tetapi, pada tahun 1937, vibrio ini menyebabkan wabah kolera yang besar dan parah di Pulau Sulawesi (Indonesia) dengan tingkat kematian lebih dari 60%. Akhirnya, pada tahun 1961, ia menjadi biang keladi pandemi ke-7, dan pada tahun 1962 pertanyaan tentang sifat koleranya akhirnya terjawab. Perbedaan antara V. cholerae dan V. eltor hanya menyangkut beberapa karakteristik. Dalam semua sifat lainnya, V. eltor pada dasarnya tidak berbeda dari V. cholerae. Selain itu, sekarang telah ditetapkan bahwa biotipe V. proteus (V.finklerpriori) mencakup seluruh kelompok vibrio, kecuali kelompok 01 (dan sekarang 0139), yang sebelumnya disebut vibrio NAG. Biotipe V. albensis diisolasi dari Sungai Elbe dan memiliki kemampuan untuk memfosforesensi, tetapi setelah kehilangannya, ia tidak berbeda dari V. proteus. Berdasarkan data tersebut, spesies Vibrio cholerae saat ini terbagi menjadi 4 biotipe, yaitu: V. cholerae 01 cholerae, V. cholerae eltor, V. cholerae 0139 bengal, dan V. cholerae non 01. Ketiga biotipe pertama tergolong dalam dua serovar, yaitu 01 dan 0139. Biovar terakhir meliputi biotipe sebelumnya, yaitu V. proteus dan V. albensis, dan diwakili oleh banyak serovar vibrio lain yang tidak diaglutinasi oleh serum 01 dan 0139, yaitu vibrio NAG.
Faktor patogenisitas vibrio kolera
 [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ]
[ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ]
Kemotaksis Vibrio cholerae
Dengan bantuan sifat-sifat ini, vibrio berinteraksi dengan sel-sel epitel. Pada mutan vibrio kolera (yang telah kehilangan kemampuan kemotaksis), virulensi berkurang secara signifikan, pada mutan Mob (yang telah kehilangan mobilitas), virulensinya menghilang sepenuhnya atau menurun tajam.
Faktor adhesi dan kolonisasi yang menyebabkan vibrio menempel pada mikrovili dan mengkolonisasi selaput lendir usus halus. Faktor adhesi meliputi musinase, hemaglutinin/protease terlarut, neuraminidase, dll. Faktor-faktor ini mendorong adhesi dan kolonisasi dengan menghancurkan zat-zat yang merupakan bagian dari lendir. Hemaglutinin/protease terlarut mendorong pemisahan vibrio dari reseptor sel epitel dan keluarnya vibrio dari usus ke lingkungan eksternal, yang memastikan penyebaran epidemiknya. Neuraminidase memperkuat ikatan antara koleragen dan sel epitel dan memfasilitasi penetrasi toksin ke dalam sel, yang meningkatkan keparahan diare.
Toksin kolera adalah koleragen.
Yang disebut racun baru yang mampu menyebabkan diare, tetapi tidak memiliki hubungan genetik atau imunologis dengan koleragen.
Faktor dermoneurotik dan hemoragik. Sifat faktor toksik ini dan perannya dalam patogenesis kolera belum diteliti secara memadai.
[ 13 ], [ 14 ], [ 15 ], [ 16 ], [ 17 ], [ 18 ]
Endotoksin dari Vibrio cholerae
Lipopolisakarida V. cholerae memiliki sifat endotoksik yang kuat dan menyebabkan keracunan umum pada tubuh.
Faktor patogenisitas utama dari vibrio cholera yang terdaftar adalah eksotoksin choleragen (CTX AB), yang menentukan patogenesis penyakit ini. Molekul kolera terdiri dari dua fragmen - A dan B. Fragmen A terdiri dari dua peptida - A1 dan A2, ia memiliki sifat khusus toksin kolera dan memberinya kualitas superantigen. Fragmen B terdiri dari 5 subunit identik. Ia melakukan dua fungsi: 1) mengenali reseptor (monosialoganglioside) enterosit dan mengikatnya; 2) membentuk saluran hidrofobik intramembran untuk lewatnya subunit A. Peptida A2 berfungsi untuk mengikat fragmen A dan B. Fungsi toksik yang sebenarnya dilakukan oleh peptida Aj (ADP-ribosiltransferase). Ia berinteraksi dengan NAD, menyebabkan hidrolisisnya; ADP-ribosa yang dihasilkan mengikat subunit pengatur adenilat siklase. Hal ini menyebabkan penghambatan hidrolisis GTP. Kompleks GTP + adenilat siklase yang dihasilkan menyebabkan hidrolisis ATP dengan pembentukan cAMP. (Jalur lain untuk akumulasi cAMP adalah penekanan oleh koleragen terhadap enzim yang menghidrolisis cAMP menjadi 5-AMP). Manifestasi fungsi gen ctxAB yang mengkode sintesis eksotoksin bergantung pada fungsi sejumlah gen patogenisitas lainnya, khususnya gen tcp (yang mengkode sintesis pili adhesi yang dikendalikan toksin - TCAP), gen pengatur toxR, toxS dan toxT, gen hap (hemaglutinin/protease terlarut) dan gen neuraminidase (neuraminidase). Oleh karena itu, kontrol genetik patogenisitas V. cholerae bersifat kompleks.
Ternyata, terdapat dua pulau patogenisitas dalam kromosom V. cholerae. Salah satunya adalah genom fag berfilamen sedang yang mengubah CTXφ, dan yang lainnya adalah genom fag berfilamen sedang yang mengubah VPIcp. Masing-masing pulau patogenisitas ini mengandung kaset gen yang ditentukan dalam profase, yang menentukan patogenisitas patogen kolera. Profaga CTXφ membawa gen CTX, gen toksin baru zot dan ace, gen ser (sintesis adhesin), dan gen ortU (sintesis produk dengan fungsi yang tidak diketahui). Kaset ini juga mencakup gen nei dan wilayah fag RS2, yang mengkode replikasi dan integrasi profag ke dalam kromosom. Gen zot, ace, dan ortU diperlukan untuk pembentukan virion fag ketika profag dikeluarkan dari kromosom patogen.
Profage VPIcp membawa gen tcp (yang mengkode produksi pili (protein TCPA)), toxT, toxR, gen act (faktor kolonisasi tambahan, gen mobilitas (integrase dan transposase)). Transkripsi gen virulensi diatur oleh tiga gen pengatur: toxR, toxS, dan toxT. Gen-gen ini secara terkoordinasi, pada tingkat transkripsi, mengubah aktivitas lebih dari 20 gen virulensi, termasuk ctxAB, tcp, dan gen lainnya. Gen pengatur utama adalah gen toxR. Kerusakan atau ketidakhadirannya menyebabkan avirulensi atau penurunan lebih dari 100 kali lipat dalam produksi toksin kolera CTX dan TCPA. Mungkin, ini adalah bagaimana ekspresi terkoordinasi gen virulensi diatur dalam pulau patogenisitas yang dibentuk oleh fag pengubah sedang dan pada spesies bakteri lainnya. Telah ditetapkan bahwa prophage K139 lain hadir dalam kromosom V. cholerae eltor, tetapi genomnya masih sedikit dipelajari.
Gen hap terlokalisasi pada kromosom. Dengan demikian, virulensi (patogenisitas) dan kapasitas epidemik V. cholerae ditentukan oleh 4 gen: ctxAB, tcp, toxR dan hap.
Berbagai metode dapat digunakan untuk mendeteksi kemampuan V. cholerae dalam menghasilkan koleragen.
Uji biologis pada kelinci. Ketika vibrio kolera disuntikkan secara intramuskular ke dalam tubuh kelinci yang sedang menyusui (berusia tidak lebih dari 2 minggu), mereka akan mengalami sindrom kolera yang umum: diare, dehidrasi, dan kematian kelinci.
Deteksi langsung koleragen dengan PCR, IFM atau reaksi hemolisis imun pasif (kolerogen mengikat Gmj eritrosit, dan eritrosit tersebut dilisiskan setelah penambahan antibodi antitoksik dan komplemen). Akan tetapi, deteksi kemampuan untuk menghasilkan toksin saja tidak cukup untuk menentukan bahaya epidemik strain tersebut. Untuk itu, perlu dideteksi keberadaan gen hap, oleh karena itu, cara terbaik dan paling dapat diandalkan untuk membedakan strain vibrio kolera toksigenik dan epidemik dari serogrup 01 dan 0139 adalah dengan PCR menggunakan primer spesifik untuk mendeteksi keempat gen patogenisitas: ctxAB, tcp, toxR dan hap.
Kemampuan V. cholerae selain serogrup 01 atau 0139 untuk menyebabkan penyakit diare sporadis atau berkelompok pada manusia mungkin disebabkan oleh keberadaan enterotoksin tipe LT atau ST, yang masing-masing merangsang sistem adenilat atau guanilat siklase, atau karena hanya ada gen ctxAB dan tidak ada gen hap.
Selama pandemi ketujuh, strain V. cholerae dengan berbagai tingkat virulensi diisolasi: kolerogenik (virulen), kolerogenik lemah (virulensi rendah), dan non-kolerogenik (non-virulen). V. cholerae non-kolerogenik, sebagai aturan, menunjukkan aktivitas hemolitik, tidak dilisiskan oleh fag diagnostik kolera HDF(5), dan tidak menyebabkan penyakit pada manusia.
Untuk pengetikan fag V. cholerae 01 (termasuk El Tor), S. Mukherjee mengusulkan beberapa set fag, yang kemudian dilengkapi dengan fag lain di Rusia. Satu set fag tersebut (1-7) memungkinkan seseorang untuk membedakan jenis fag di antara V. cholerae 0116. Untuk identifikasi V. cholerae El Tor yang toksigenik dan non-toksigenik, alih-alih HDF-3, HDF-4 dan HDF-5, fag CTX* (melisis vibrio El Tor yang toksigenik) dan CTX" (melisis vibrio El Tor yang non-toksigenik) kini diusulkan di Rusia.
Resistensi terhadap patogen kolera
Vibrio kolera bertahan hidup dengan baik pada suhu rendah; mereka tetap layak dalam es hingga 1 bulan; dalam air laut - hingga 47 hari, dalam air sungai - dari 3-5 hari hingga beberapa minggu, dalam air mineral mendidih mereka bertahan hidup selama lebih dari 1 tahun, dalam tanah - dari 8 hari hingga 3 bulan, dalam tinja segar - hingga 3 hari, pada produk yang direbus (nasi, mie, daging, bubur, dll.) mereka bertahan hidup selama 2-5 hari, pada sayuran mentah - 2-4 hari, pada buah-buahan - 1-2 hari, dalam susu dan produk susu - 5 hari; ketika disimpan dalam dingin, masa bertahan hidup meningkat 1-3 hari; pada linen linen yang terkontaminasi dengan tinja, mereka bertahan hidup hingga 2 hari, dan pada bahan lembab - seminggu. Vibrio kolera mati dalam 5 menit pada suhu 80 °C, dan langsung pada 100 °C; mereka sangat sensitif terhadap asam; mereka mati dalam 5-15 menit di bawah pengaruh kloramin dan desinfektan lainnya. Bakteri ini sensitif terhadap pengeringan dan sinar matahari langsung, tetapi dapat bertahan hidup dengan baik dan dalam waktu lama, bahkan berkembang biak di perairan terbuka dan air limbah yang kaya akan bahan organik, dengan pH basa dan suhu di atas 10-12 °C. Bakteri ini sangat sensitif terhadap klorin: dosis klorin aktif 0,3-0,4 mg/l air dalam 30 menit menyebabkan disinfeksi yang andal dari vibrio kolera.
Vibrio yang bersifat patogen terhadap manusia yang tidak termasuk dalam spesies Vibrio Cholerae
Genus Vibrio mencakup lebih dari 25 spesies, yang, selain V. cholerae, setidaknya delapan mampu menyebabkan penyakit pada manusia: V. parahaemolyticus, V. alginolyticus, V. vulnificus, V. fluvialis, V. fumissii, V. mimicus, V. damsela, dan V. hollisae. Semua vibrio ini menghuni laut dan teluk. Infeksi terjadi baik melalui berenang atau melalui makan makanan laut. Telah ditemukan bahwa vibrio kolera dan non-kolera dapat menyebabkan tidak hanya gastroenteritis, tetapi juga infeksi luka. Kemampuan ini telah ditemukan dalam kelompok V. cholerae 01 dan non-01, V. parahaemolyticus, V. alginolyticus, V. mimicus, V. damsela, dan V. vulnificus. Mereka menyebabkan proses inflamasi pada jaringan lunak ketika mereka rusak oleh cangkang hewan laut atau ketika bersentuhan langsung dengan air laut yang terinfeksi.
Dari vibrio patogen non-kolera yang terdaftar, yang paling menarik secara praktis adalah V. parahaemolyticus, V. alginolyticus, V. vulnificus dan V. fluvialis.
V. parahaemolyticus - vibrio parahaemolytic - pertama kali diisolasi di Jepang pada tahun 1950 selama wabah besar keracunan makanan yang disebabkan oleh konsumsi sarden semi-kering (kematian adalah 7,5%). Agen penyebab termasuk dalam genus Vibrio oleh R. Sakazaki pada tahun 1963. Ia membagi strain yang dipelajari menjadi 2 spesies: V. parahaemolyticus dan V. alginolyticus. Kedua spesies ditemukan di air laut pesisir dan di penghuninya, mereka adalah halofil (bahasa Yunani hals - garam); tidak seperti vibrio biasa, vibrio halofilik tidak tumbuh pada media tanpa NaCl dan bereproduksi dengan baik pada konsentrasi tinggi. Afiliasi spesies vibrio halofilik ditentukan oleh kemampuannya untuk memfermentasi sukrosa, membentuk asetilmetilkarbinol, dan bereproduksi dalam PV dengan 10% NaCl. Semua fitur ini melekat pada spesies V. alginolyticus, tetapi tidak ada pada V. parahaemolyticus.
Vibrio parahaemolyticus memiliki tiga jenis antigen: antigen flagellar H yang labil terhadap panas, antigen O yang stabil terhadap panas yang tidak hancur dengan pemanasan hingga 120 °C selama 2 jam, dan antigen K permukaan yang hancur dengan pemanasan. Kultur V. parahaemolyticus yang baru diisolasi memiliki antigen K yang terdefinisi dengan baik yang melindungi vibrio hidup dari aglutinasi oleh serum O homolog. Antigen H sama untuk semua galur, tetapi antigen H dari monotrichus berbeda dari antigen H dari peritrich. Menurut antigen O, V. parahaemolyticus dibagi menjadi 14 serogroup. Dalam serogroup, vibrio dibagi menjadi serotipe menurut antigen K, yang jumlah totalnya adalah 61. Skema antigenik V. parahaemolyticus telah dikembangkan untuk galurnya yang diisolasi dari manusia saja.
Patogenisitas V. parahaemolyticus dikaitkan dengan kemampuannya untuk mensintesis hemolisin, yang memiliki sifat enterotoksik. Yang terakhir dideteksi menggunakan metode Kanagawa. Esensinya terletak pada kenyataan bahwa V. parahaemolyticus, patogen bagi manusia, menyebabkan hemolisis yang jelas pada agar darah yang mengandung 7% NaCl. Pada agar darah yang mengandung kurang dari 5% NaCl, hemolisis disebabkan oleh banyak strain V. parahaemolyticus, dan pada agar darah dengan 7% NaCl - hanya strain dengan sifat enteropatogenik. Vibrio parahaemolyticus ditemukan di pantai Jepang, Kaspia, Hitam dan laut lainnya. Ini menyebabkan infeksi toksik bawaan makanan dan penyakit seperti disentri. Infeksi terjadi ketika memakan makanan laut mentah atau setengah mentah yang terinfeksi V- parahaemolyticus (ikan laut, tiram, krustasea, dll.).
Di antara delapan vibrio non-kolera yang disebutkan di atas, yang paling patogen bagi manusia adalah V. vulnificus, yang pertama kali dideskripsikan pada tahun 1976 sebagai Beneckea vulnificus dan kemudian direklasifikasi sebagai Vibrio vulnificus pada tahun 1980. Bakteri ini sering ditemukan di air laut dan penghuninya serta menyebabkan berbagai penyakit pada manusia. Strain V. vulnificus yang berasal dari laut dan klinis tidak berbeda satu sama lain baik secara fenotip maupun genetik.
Infeksi luka yang disebabkan oleh V. vulnificus berkembang dengan cepat dan menyebabkan terbentuknya tumor dengan nekrosis jaringan berikutnya, disertai demam, menggigil, terkadang nyeri hebat, dan dalam beberapa kasus memerlukan amputasi.
V. vulnificus diketahui menghasilkan eksotoksin. Percobaan pada hewan menunjukkan bahwa patogen tersebut menyebabkan kerusakan lokal yang parah dengan perkembangan edema dan nekrosis jaringan, diikuti oleh kematian. Peran eksotoksin dalam patogenesis penyakit ini sedang dipelajari.
Selain infeksi luka, V. vulnificus dapat menyebabkan pneumonia pada korban tenggelam dan endometritis pada wanita setelah terpapar air laut. Bentuk infeksi paling parah yang disebabkan oleh V. vulnificus adalah septikemia primer yang terkait dengan konsumsi tiram mentah (dan mungkin hewan laut lainnya). Penyakit ini berkembang sangat cepat: pasien mengalami malaise, demam, menggigil, dan lemas, kemudian hipotensi berat, yang merupakan penyebab utama kematian (angka kematian sekitar 50%).
V. fluvialis pertama kali dideskripsikan sebagai patogen gastroenteritis pada tahun 1981. Bakteri ini termasuk dalam subkelompok vibrio patogen non-kolera yang memiliki arginin dihidrolase tetapi tidak memiliki netornitin dan lisin dekarboksilase (V. fluvialis, V. furnissii, V. damsela, yaitu, secara fenotip mirip dengan Aeromonas). V. fluvialis merupakan agen penyebab gastroenteritis yang umum, yang disertai dengan muntah hebat, diare, nyeri perut, demam, dan dehidrasi berat atau sedang. Faktor patogenik utamanya adalah enterotoksin.
Epidemiologi Kolera
Sumber utama infeksi hanya orang - pasien dengan kolera atau pembawa vibrio, serta air yang terkontaminasi dengannya. Tidak ada hewan di alam yang terkena kolera. Rute infeksi adalah feco-oral. Rute infeksi: a) yang utama - melalui air yang digunakan untuk minum, mandi dan kebutuhan rumah tangga; b) kontak-rumah tangga dan c) melalui makanan. Semua epidemi dan pandemi kolera utama dikaitkan dengan air. Vibrio kolera memiliki mekanisme adaptif yang memastikan keberadaan populasi mereka baik dalam tubuh manusia maupun di ekosistem tertentu di badan air terbuka. Diare parah, yang disebabkan oleh vibrio kolera, menyebabkan pembersihan usus dari bakteri yang bersaing dan berkontribusi pada penyebaran patogen yang meluas di lingkungan, terutama di air limbah dan di badan air terbuka tempat mereka dibuang. Seseorang yang terkena kolera mengeluarkan patogen dalam jumlah yang sangat besar - dari 100 juta hingga 1 miliar per 1 ml feses, pembawa vibrio mengeluarkan 100-100.000 vibrio dalam 1 ml, dosis infeksiusnya sekitar 1 juta vibrio. Durasi ekskresi vibrio kolera pada pembawa yang sehat adalah 7 hingga 42 hari dan 7-10 hari pada mereka yang telah pulih. Ekskresi yang lebih lama sangat jarang terjadi.
Keunikan kolera adalah bahwa setelahnya, sebagai aturan, tidak ada pembawa jangka panjang dan tidak ada fokus endemik yang stabil terbentuk. Namun, seperti yang telah ditunjukkan di atas, karena pencemaran badan air terbuka dengan air limbah yang mengandung sejumlah besar zat organik, deterjen, dan garam dapur, di musim panas vibrio kolera tidak hanya bertahan di dalamnya untuk waktu yang lama, tetapi bahkan berkembang biak.
Yang sangat penting secara epidemiologis adalah fakta bahwa vibrio kolera dari kelompok 01, baik yang non-toksikogenik maupun toksigenik, dapat bertahan lama di berbagai ekosistem perairan sebagai bentuk yang tidak dibudidayakan. Dengan menggunakan reaksi berantai polimerase, gen vct dari bentuk V. chokrae yang tidak dibudidayakan dideteksi di berbagai badan air di sejumlah wilayah endemik CIS selama studi bakteriologis negatif.
Fokus endemik vibrio kolera El Tor adalah Indonesia, munculnya biang keladi pandemi ketujuh ini dari sana diyakini terkait dengan meluasnya hubungan ekonomi Indonesia dengan dunia luar setelah merdeka, dan durasi serta perkembangan pandemi yang begitu cepat, terutama gelombang keduanya, secara menentukan dipengaruhi oleh kurangnya kekebalan terhadap kolera dan berbagai pergolakan sosial di negara-negara Asia, Afrika, dan Amerika.
Dalam kasus kolera, serangkaian tindakan anti-epidemi diambil, di antaranya yang utama dan menentukan adalah deteksi dan isolasi aktif dan tepat waktu (rawat inap, perawatan) pasien dalam bentuk akut dan atipikal dan pembawa vibrio yang sehat; tindakan diambil untuk mencegah kemungkinan rute infeksi; perhatian khusus diberikan pada pasokan air (klorinasi air minum), kepatuhan terhadap kondisi sanitasi dan higienis di perusahaan makanan, di lembaga anak-anak, tempat umum; kontrol ketat, termasuk bakteriologis, dilakukan terhadap badan air terbuka, imunisasi populasi dilakukan, dll.
Gejala Kolera
Masa inkubasi kolera bervariasi dari beberapa jam hingga 6 hari, paling sering 2-3 hari. Setelah memasuki lumen usus halus, vibrio kolera, karena mobilitas dan kemotaksisnya terhadap selaput lendir, diarahkan ke lendir. Untuk menembusnya, vibrio menghasilkan sejumlah enzim: neuraminidase, mukinase, protease, lesitinase, yang menghancurkan zat-zat yang terkandung dalam lendir dan memfasilitasi pergerakan vibrio ke sel-sel epitel. Dengan adhesi, vibrio menempel pada glikokaliks epitel dan, kehilangan mobilitas, mulai berkembang biak secara intensif, menjajah mikrovili usus halus (lihat sisipan berwarna, Gambar 101.2), dan secara bersamaan menghasilkan sejumlah besar eksotoksin-kolerogen. Molekul koleragen mengikat monosialoganglioside Gni! Dan menembus membran sel, di mana mereka mengaktifkan sistem adenilat siklase, dan akumulasi cAMP menyebabkan hipersekresi cairan, kation dan anion Na, HCO, Kl, Cl dari enterosit, yang menyebabkan diare kolera, dehidrasi dan desalinasi tubuh. Ada tiga jenis penyakit:
- penyakit diare yang parah dan menyebabkan dehidrasi yang menyebabkan kematian pasien dalam beberapa jam;
- perjalanan penyakit yang kurang parah, atau diare tanpa dehidrasi;
- perjalanan penyakit yang asimtomatik (pembawa vibrio).
Dalam kasus kolera yang parah, pasien mengalami diare, frekuensi tinja meningkat, tinja menjadi lebih banyak, menjadi encer, kehilangan bau tinja dan tampak seperti kaldu beras (cairan keruh dengan residu lendir dan sel epitel yang mengambang di dalamnya). Kemudian muntah yang melemahkan terjadi, pertama-tama dari isi usus, dan kemudian muntahan tersebut tampak seperti kaldu beras. Suhu pasien turun di bawah normal, kulit menjadi kebiru-biruan, keriput dan dingin - kolera algid. Sebagai akibat dehidrasi, darah mengental, sianosis berkembang, kelaparan oksigen, fungsi ginjal menurun tajam, kejang muncul, pasien kehilangan kesadaran dan kematian terjadi. Angka kematian akibat kolera selama pandemi ketujuh bervariasi dari 1,5% di negara-negara maju hingga 50% di negara-negara berkembang.
Kekebalan pasca infeksi bersifat kuat, bertahan lama, dan penyakit yang kambuh jarang terjadi. Kekebalan bersifat antitoksik dan antimikroba, disebabkan oleh antibodi (antitoksin bertahan lebih lama daripada antibodi antimikroba), sel memori imun, dan fagosit.
Diagnostik laboratorium kolera
Metode utama dan penentu untuk mendiagnosis kolera adalah bakteriologis. Bahan untuk pemeriksaan dari pasien adalah feses dan muntahan; feses diperiksa untuk mengetahui adanya vibrio; bagian usus halus dan kantong empedu yang diligasi diambil untuk pemeriksaan dari orang yang meninggal karena kolera; dari benda-benda di lingkungan luar, air dari waduk terbuka dan air limbah paling sering diperiksa.
Ketika melakukan studi bakteriologis, tiga kondisi berikut harus dipenuhi:
- menabur bahan dari pasien secepat mungkin (vibrio kolera bertahan hidup dalam tinja untuk jangka waktu yang singkat);
- wadah tempat pengambilan bahan tidak boleh didisinfeksi dengan bahan kimia dan tidak boleh mengandung jejak bahan kimia, karena vibrio kolera sangat sensitif terhadap bahan kimia;
- menghilangkan kemungkinan kontaminasi dan infeksi pada orang lain.
Kultur diisolasi menurut skema berikut: disemai pada PV, secara bersamaan pada MPA alkali atau media selektif apa pun (TCBS adalah yang terbaik). Setelah 6 jam, film yang terbentuk pada PV diperiksa, dan jika perlu, pemindahan dilakukan ke PV kedua (laju penyemaian vibrio kolera dalam kasus ini meningkat sebesar 10%). Dari PV, pemindahan dilakukan ke MPA alkali. Koloni yang mencurigakan (transparan-kaca) dipindahkan untuk mendapatkan kultur murni, yang diidentifikasi berdasarkan sifat morfologi, kultur, biokimia, motilitas dan akhirnya diketik menggunakan serum aglutinasi diagnostik O-, OR-, Inaba dan Ogawa dan fag (HDF). Berbagai pilihan untuk diagnostik yang dipercepat telah diusulkan, yang terbaik adalah metode serologis luminescent. Ini memungkinkan untuk mendeteksi vibrio kolera secara langsung dalam bahan uji (atau setelah kultivasi awal dalam dua tabung reaksi dengan 1% PV, yang salah satunya ditambahkan fag kolera) dalam waktu 1,5-2 jam. Untuk deteksi cepat kolera vibrio, IEM Nizhny Novgorod telah mengusulkan seperangkat cakram indikator kertas yang terdiri dari 13 uji biokimia (oksidase, indol, urease, laktosa, glukosa, sukrosa, manosa, arabinosa, manitol, inositol, arginina, ornitina, lisin), yang memungkinkan untuk membedakan perwakilan genus Vibrio dari genus Aeromonas, Plesiomonas, Pseudomonas, Comamonas dan dari famili Enterobacteriaceae. Untuk deteksi cepat kolera vibrio dalam tinja dan benda-benda lingkungan, RPGA dengan antibodi diagnosticum dapat digunakan. Untuk mendeteksi bentuk kolera vibrio yang tidak dibudidayakan dalam benda-benda lingkungan, hanya metode reaksi berantai polimerase yang digunakan.
Dalam kasus di mana V. cholerae non-Ol-group diisolasi, mereka harus diketik menggunakan serum aglutinasi yang sesuai dari serogrup lain. Isolasi V. cholerae non-Ol-group dari pasien diare (termasuk diare mirip kolera) memerlukan tindakan anti-epidemi yang sama seperti dalam kasus isolasi V. cholerae Ol-group. Jika perlu, keberadaan gen patogenisitas ctxAB, tcp, toxR dan hap ditentukan dalam vibrio tersebut menggunakan PCR.
Diagnosis serologis kolera bersifat tambahan. Untuk tujuan ini, reaksi aglutinasi dapat digunakan, tetapi lebih baik untuk menentukan titer antibodi vibriosidal atau antitoksin (antibodi terhadap kolera ditentukan dengan metode enzim immunoassay atau immunofluorescence).
Diagnostik laboratorium vibrio patogen non-kolera
Metode utama untuk mendiagnosis penyakit yang disebabkan oleh vibrio patogen non-kolera adalah bakteriologis dengan menggunakan media selektif seperti TCBS, MacConkey, dll. Kepemilikan kultur yang diisolasi ke dalam genus Vibrio ditentukan berdasarkan karakteristik utama bakteri dari genus ini.
Pengobatan Kolera
Pengobatan pasien kolera terutama harus terdiri dari rehidrasi dan pemulihan metabolisme air-garam normal. Untuk tujuan ini, disarankan untuk menggunakan larutan garam, misalnya, dengan komposisi berikut: NaCl - 3,5; NaHC03 - 2,5; KCl - 1,5 dan glukosa - 20,0 g per 1 liter air. Pengobatan yang didukung secara patogenetik tersebut dikombinasikan dengan terapi antibiotik rasional memungkinkan untuk mengurangi angka kematian pada kolera hingga 1% atau kurang.
Pencegahan khusus kolera
Untuk menciptakan kekebalan buatan, diusulkan vaksin kolera, termasuk vaksin yang dibuat dari strain Inaba dan Ogawa yang telah dimatikan; toksoid kolera untuk penggunaan subkutan dan vaksin bivalen kimia enteral yang terdiri dari anatoksin dan antigen somatik dari serotipe Inaba dan Ogawa, karena kekebalan silang tidak terbentuk. Namun, durasi kekebalan pasca vaksinasi tidak lebih dari 6-8 bulan, sehingga vaksinasi hanya dilakukan sesuai indikasi epidemiologis. Profilaksis antibiotik telah terbukti baik pada fokus kolera, khususnya tetrasiklin, yang sangat sensitif terhadap vibrio kolera. Antibiotik lain yang efektif terhadap V. cholerae dapat digunakan untuk tujuan yang sama.