
Sintesis, sekresi dan metabolisme katekolamin
Terakhir ditinjau: 19.10.2021

Semua konten iLive ditinjau secara medis atau diperiksa fakta untuk memastikan akurasi faktual sebanyak mungkin.
Kami memiliki panduan sumber yang ketat dan hanya menautkan ke situs media terkemuka, lembaga penelitian akademik, dan, jika mungkin, studi yang ditinjau secara medis oleh rekan sejawat. Perhatikan bahwa angka dalam tanda kurung ([1], [2], dll.) Adalah tautan yang dapat diklik untuk studi ini.
Jika Anda merasa salah satu konten kami tidak akurat, ketinggalan zaman, atau dipertanyakan, pilih dan tekan Ctrl + Enter.
Lapisan serebral kelenjar adrenal menghasilkan senyawa yang jauh dari steroid dari struktur. Mereka mengandung inti 3,4-dihydroxyphenyl (catechol) dan disebut katekolamin. Ini termasuk adrenalin, norepinefrin dan dopamin beta-oksitiramin.
Urutan sintesis katekolamin cukup sederhana: tirosin → dioksifenilalanin (DOPA) → dopamin → noradrenalin → adrenalin. Tirosin memasuki tubuh dengan makanan, tetapi juga dapat terbentuk dari fenilalanin di hati di bawah aksi fenilalanin hidroksilase. Produk akhir dari transformasi tirosin dalam jaringan berbeda. Di medula adrenal proses hasil untuk melangkah pembentukan adrenalin, di ujung saraf simpatis - noradrenalin, dalam neuron tertentu dari sistem katekolamin saraf sintesis dopamin formulir yang telah dilengkapi pusat.
Konversi tirosin ke DOPA dikatalisis oleh tirosin hidroksilase, kofaktor diantaranya adalah tetrahidro-biopterin dan oksigen. Diyakini bahwa enzim inilah yang membatasi kecepatan seluruh proses biosintesis katekolamin dan dihambat oleh produk akhir dari proses. Tirosin hidroksilase adalah objek utama efek regulasi pada biosintesis katekolamin.
Dopa konversi dopamin dikatalisis oleh enzim dopa-dekarboksilase (kofaktor - piridoksal) yang relatif non-spesifik dan dekarboksilasi, dan lainnya aromatik asam L-amino. Namun, ada indikasi kemungkinan memodifikasi sintesis katekolamin dengan mengubah aktivitas dan enzim ini. Dalam beberapa neuron tidak ada enzim untuk konversi dopamin lebih lanjut, dan itu adalah produk akhir. Jaringan lain mengandung dopamin-beta-hidroksilase (kofaktor adalah tembaga, asam askorbat dan oksigen), yang mengubah dopamin menjadi noradrenalin. Di medula adrenal (tetapi tidak di ujung saraf simpatetik) ada phenylethanolamine - methyltransferase, yang menghasilkan adrenalin dari norepinefrin. Dalam hal ini S-adenosylmethionine berfungsi sebagai donor dari kelompok metil.
Penting untuk diingat bahwa sintesis phenylethanolamine-N-Metiltransferazy diinduksi oleh glukokortikoid yang jatuh di lapisan korteks serebral dari sistem vena porta. Hal ini mungkin menjelaskan kebohongan fakta menggabungkan dua kelenjar endokrin yang berbeda dalam satu tubuh. Arti sintesis glukokortikoid adrenalin ditekankan oleh fakta bahwa sel-sel medula adrenal memproduksi norepinephrine, yang disusun di sekitar pembuluh arteri, sementara sel-sel darah diperoleh adrenalinprodutsiruyuschie dasarnya sinus vena, terlokalisasi di korteks adrenal.
Runtuhnya katekolamin terjadi terutama di bawah pengaruh dua sistem enzim: catechol-O-methyltransferase (COMT) dan monoamine oxidase (MAO). Cara utama epinefrin dan norepinefrin dari pembusukan ditunjukkan secara skematis pada Gambar. 54. Dalam aksi COMT di hadapan donor katekolamin metil kelompok S-adrenozilmetionina dan diubah menjadi normetanephrine metanephrine (3-O-metil-turunan dari epinefrin dan norepinefrin), yang di bawah pengaruh MAO diubah menjadi aldehida dan lebih (dengan adanya aldehida) di vanillyl-mandelic acid (ICH) - produk degradasi utama noradrenalin dan adrenalin. Dalam kasus yang sama, ketika pertama kali terkena tindakan katekolamin MAO, tidak COMT, mereka dikonversi ke aldehida 3,4-dioksimindalevy, dan kemudian di bawah pengaruh aldehida dan COMT - asam 3,4-dioksimindalnuyu dan IUD. Di hadapan alkohol dehidrogenase katekolamin dapat membentuk 3-metoksi-4-oksifenilglikol, akhir-produk utama degradasi epinefrin dan norepinefrin dalam SSP.
Disintegrasi dopamin hasil sama, kecuali bahwa metabolitnya yang tanpa gugus hidroksil pada atom beta-karbon, dan karena itu, bukannya asam vanillyl-mandelic dibentuk homovanillic (HVA) dan asam 3-metoksi-4-oksifeniluksusnaya.
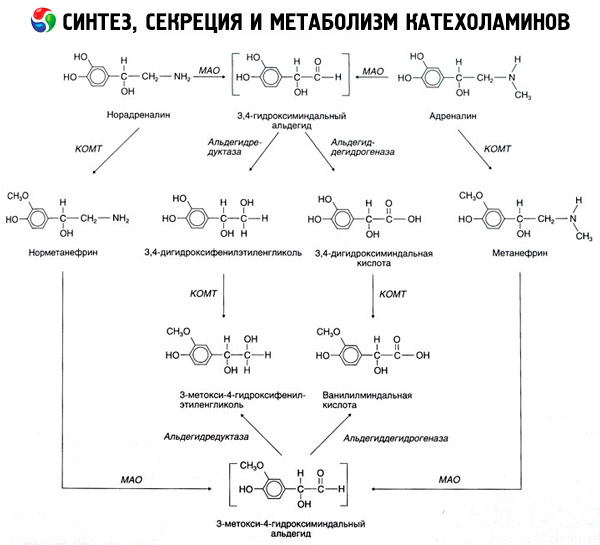
Adanya jalur quinoid untuk oksidasi molekul katekolamin, di mana produk antara dengan aktivitas biologis diucapkan, juga dapat dipostulasikan.
Dibentuk oleh aksi enzim sitosol, adrenalin dan noradrenalin di ujung saraf simpatis, medula adrenal dan masukkan butiran sekretori yang melindungi mereka dari aksi enzim degradatif. Penangkapan katekolamin dengan butiran membutuhkan biaya energi. Dalam butiran chromaffin dari katekolamin medula adrenal tegas terikat ATP (dalam rasio 4: 1) dan protein spesifik - chromogranin yang mencegah difusi hormon dari butiran dalam sitoplasma.
Stimulus langsung untuk sekresi katekolamin tampaknya penetrasi sel kalsium merangsang eksositosis (butiran fusi membran dengan permukaan sel dan kesenjangan mereka dengan hasil keseluruhan konten larut - katekolamin, dopamin-beta-hidroksilase, ATP dan chromogranin - ke dalam cairan ekstraseluler) .
Efek fisiologis katekolamin dan mekanisme aksi mereka
Efek katekolamin dimulai dengan interaksi dengan reseptor spesifik sel target. Jika reseptor tiroid dan hormon steroid terlokalisasi dalam sel, reseptor katekolamin (serta asetilkolin dan peptida hormon) yang hadir pada permukaan sel luar.
Ini telah lama ditetapkan bahwa sehubungan beberapa reaksi adrenalin atau noradrenalin lebih efektif daripada katekolamin isoproterenol sintetis, sedangkan untuk orang lain efek lebih unggul aksi isoproterenol epinefrin atau norepinefrin. Atas dasar ini, konsep dikembangkan untuk kehadiran dalam jaringan dua jenis adrenoreseptor: alfa dan beta, dan di beberapa dari mereka hanya satu dari dua jenis ini yang dapat hadir. Isoproterenol adalah agonis beta adrenoreseptor yang paling kuat, sedangkan senyawa sintetis phenylephrine adalah agonis alpha-adrenoreseptor yang paling kuat. Katekolamin alami - adrenalin dan noradrenalin - dapat berinteraksi dengan reseptor dari kedua jenis, tetapi adrenalin mengambil afinitas yang lebih besar untuk versi beta, dan norepinefrin - alpha-reseptor.
Katekolamin kuat mengaktifkan reseptor beta-adrenergik jantung daripada reseptor beta dari otot polos, yang memungkinkan beta-jenis dibagi menjadi subtipe: beta1-reseptor (jantung, sel-sel lemak) dan reseptor beta2 (bronkus, pembuluh darah, dll ...). Aksi isoproterenol pada tindakan unggul reseptor beta1 adrenalin dan noradrenalin hanya 10 kali, sedangkan beta2-reseptor bertindak 100-1000 kali lebih kuat dari katekolamin alami.
Penggunaan antagonis spesifik (phentolamine dan phenoxybenzamine terhadap alpha dan propranolol untuk reseptor beta) mengkonfirmasi kecukupan klasifikasi adrenoreseptor. Dopamin mampu berinteraksi dengan baik alfa dan beta-reseptor, tetapi dalam berbagai jaringan (otak, kelenjar pituitari, kapal) ditemukan dan reseptor dopaminergik sendiri blocker tertentu yang haloperidol. Jumlah reseptor beta berkisar dari 1000 hingga 2000 per sel. Efek biologis katekolamin dimediasi oleh reseptor beta terkait, sebagai suatu peraturan, dengan aktivasi adenilat siklase dan peningkatan kandungan intraseluler cAMP. Reseptor dan enzim, meskipun mereka terhubung secara fungsional, tetapi mewakili makromolekul yang berbeda. Dalam modulasi aktivitas adenilat siklase, di bawah pengaruh kompleks hormon-reseptor, guanosin trifosfat (GTP) dan nukleotida purin lainnya terlibat. Dengan meningkatkan aktivitas enzim, mereka tampaknya mengurangi afinitas reseptor beta untuk agonis.
Fenomena peningkatan kepekaan struktur denervasi telah lama diketahui. Sebaliknya, paparan agonis yang berkepanjangan mengurangi sensitivitas jaringan target. Studi tentang reseptor beta diizinkan untuk menjelaskan fenomena ini. Itu menunjukkan bahwa tindakan isoproterenol yang berkepanjangan menyebabkan hilangnya kepekaan adenilat siklase karena penurunan jumlah reseptor beta.
Proses desensitisasi tidak memerlukan aktivasi sintesis protein dan mungkin karena pembentukan bertahap kompleks reseptor hormon ireversibel. Sebaliknya, pengenalan 6-oxydopamine, menghancurkan ujung simpatetik, disertai dengan peningkatan jumlah reseptor beta reaktif dalam jaringan. Tidak dikecualikan bahwa peningkatan aktivitas saraf simpatis menentukan desensitisasi terkait usia pembuluh darah dan jaringan lemak dalam kaitannya dengan katekolamin.
Jumlah adrenoreseptor di organ yang berbeda dapat dikontrol oleh hormon lain. Dengan demikian, estradiol meningkat, dan progesteron mengurangi jumlah reseptor alfa-adrenergik di uterus, yang disertai dengan peningkatan dan penurunan yang sesuai dalam respons kontraktalnya terhadap katekolamin. Jika "utusan kedua" intraseluler yang dibentuk oleh aksi agonis β-reseptor tentu cAMP, maka kasus pemancar pengaruh alpha-adrenergik lebih rumit. Diasumsikan bahwa ada berbagai mekanisme: penurunan tingkat cAMP, peningkatan isi cAMP, modulasi dinamika seluler kalsium,
Untuk mereproduksi berbagai efek dalam tubuh, dosis epinefrin, yang 5-10 kali lebih rendah dari norepinefrin, biasanya diperlukan. Meskipun yang terakhir lebih efektif untuk reseptor a dan beta1-adrenergik, penting untuk diingat bahwa kedua katekolamin endogen dapat berinteraksi dengan reseptor alfa dan beta. Oleh karena itu, respon biologis dari tubuh ini untuk aktivasi adrenergik sangat tergantung pada jenis reseptor yang ada di dalamnya. Namun, ini tidak berarti bahwa aktivasi selektif dari saraf atau humoral dari sistem simpatik-adrenal tidak mungkin. Dalam banyak kasus ada aktivitas intensif dari berbagai tautannya. Dengan demikian, diasumsikan bahwa mengaktifkan refleks hipoglikemia medula adrenal, sedangkan penurunan tekanan darah (hipotensi postural) disertai pelepasan terutama norepinefrin dari ujung saraf simpatik.
Adrenoreseptor dan efek aktivasi mereka di berbagai jaringan
|
Sistem, organ |
Jenis adrenoceptor |
Reaksi |
|
Sistem kardiovaskular: |
||
|
Hati |
Beta |
Peningkatan frekuensi kontraksi, konduksi dan kontraktilitas |
|
Arterioles: |
||
|
Kulit dan selaput lendir |
Alpha |
Pengurangan |
|
Otot rangka |
Beta |
Pengurangan Ekstensi |
|
Organ perut |
Alpha (selengkapnya) |
Pengurangan |
|
Beta |
Ekstensi |
|
|
Vena |
Alpha |
Pengurangan |
|
Sistem pernapasan: |
||
|
Otot-otot bronkus |
Beta |
Ekstensi |
|
Sistem pencernaan: |
||
|
Perut |
Beta |
Fungsi motorik menurun |
|
Usus |
Alpha |
Pengurangan sfingter |
|
Limpa |
Alpha |
Pengurangan |
|
Beta |
Relaksasi |
|
|
Bagian rahasia pankreas secara eksternal |
Alpha |
Sekresi menurun |
|
Sistem Genitourinary: |
Alpha |
Pengurangan sfingter |
|
Kandung kemih |
Beta |
Santai otot pengusir setan |
|
Organ seksual laki-laki |
Alpha |
Ejakulasi |
|
Mata |
Alpha |
Pupil melebar |
|
Kulit |
Alpha |
Meningkat berkeringat |
|
Kelenjar ludah |
Alpha |
Isolasi potasium dan air |
|
Beta |
Sekresi amilase |
|
|
Kelenjar endokrin: |
||
|
Pulau pankreas |
||
|
Sel beta |
Alpha (selengkapnya) |
Penurunan sekresi insulin |
|
Beta |
Peningkatan sekresi insulin |
|
|
Sel alfa |
Beta |
Peningkatan sekresi glukagon |
|
8-sel |
Beta |
Peningkatan sekresi somatostatin |
|
Hipotalamus dan hipofisis: |
||
|
Somatotrof |
Alpha |
Peningkatan sekresi STH |
|
Beta |
Pengurangan sekresi STH |
|
|
Laktotrof |
Alpha |
Penurunan sekresi prolaktin |
|
Tirotrof |
Alpha |
Penurunan sekresi TSH |
|
Corticotrophs |
Alpha |
Peningkatan sekresi ACTH |
| beta | Penurunan sekresi ACTH | |
|
Kelenjar tiroid: |
||
|
Sel-sel folikel |
Alpha |
Penurunan sekresi tiroksin |
|
Beta |
Peningkatan sekresi tiroksin |
|
|
Sel parafollicular (K) |
Beta |
Peningkatan sekresi kalsitonin |
|
Kelenjar paratiroid |
Beta |
Peningkatan sekresi PTH |
|
Ginjal |
Beta |
Meningkatnya sekresi renin |
|
Perut |
Beta |
Meningkatkan sekresi gastrin |
|
Pertukaran dasar |
Beta |
Peningkatan konsumsi oksigen |
|
Hati |
? |
Peningkatan glikogenolisis dan glukoneogenesis dari hasil glukosa; meningkatkan ketogenesis dengan pelepasan tubuh keton |
|
Jaringan adiposa |
Beta |
Peningkatan lipolisis dengan pelepasan asam lemak bebas dan gliserol |
|
Otot rangka |
Beta |
Peningkatan glikolisis dengan pelepasan piruvat dan laktat; penurunan proteolisis dengan penurunan hasil alanin, glutamin |
Hal ini penting untuk diingat bahwa hasil pemberian intravena katekolamin tidak selalu cukup mencerminkan efek dari senyawa endogen. Hal ini berlaku terutama untuk noradrenalin, karena berdiri dalam tubuh sebagian besar tidak darah, tetapi langsung di celah sinaps. Oleh karena itu endogen norepinefrin mengaktifkan, misalnya, tidak hanya reseptor alfa vaskular (tekanan darah meningkat) tetapi juga beta-reseptor jantung (palpitasi), sedangkan administrasi lead eksternal noradrenalin terutama untuk aktivasi reseptor alpha pembuluh darah dan refleks (melalui vagus) perlambatan palpitasi.
Dosis rendah epinefrin mengaktifkan terutama reseptor beta dari pembuluh otot dan jantung, mengakibatkan penurunan resistensi pembuluh darah perifer dan meningkatkan volume jantung. Dalam beberapa kasus, efek pertama mungkin mendominasi, dan setelah pemberian adrenalin, hipotensi berkembang. Dalam dosis yang lebih tinggi, adrenalin juga mengaktifkan reseptor alfa, yang disertai dengan peningkatan resistensi pembuluh darah perifer dan dengan latar belakang peningkatan volume menit jantung mengarah ke peningkatan tekanan darah. Namun, efeknya pada reseptor beta vaskular juga dipertahankan. Akibatnya, peningkatan tekanan sistolik melebihi nilai yang sama dari tekanan diastolik (peningkatan tekanan nadi). Dengan diperkenalkannya dosis yang lebih besar, efek alpha-mimetik dari epinefrin mulai berlaku: peningkatan sistolik dan diastolik meningkat secara paralel, keduanya di bawah pengaruh noradrenalin.
Efek katekolamin pada metabolisme terdiri dari efek langsung dan tidak langsung mereka. Yang pertama diwujudkan terutama melalui reseptor beta. Proses yang lebih kompleks terkait dengan hati. Meskipun peningkatan glikogenolisis hati secara tradisional telah dianggap sebagai hasil dari aktivasi beta-reseptor, ada juga data tentang keterlibatan reseptor alfa dalam hal ini. Efek dimediasi katekolamin berhubungan dengan modulasi sekresi banyak hormon lain, misalnya insulin. Dalam aksi adrenalin pada sekresinya, komponen alpha-adrenergik jelas mendominasi, karena ini menunjukkan bahwa setiap stres disertai dengan penghambatan sekresi insulin.
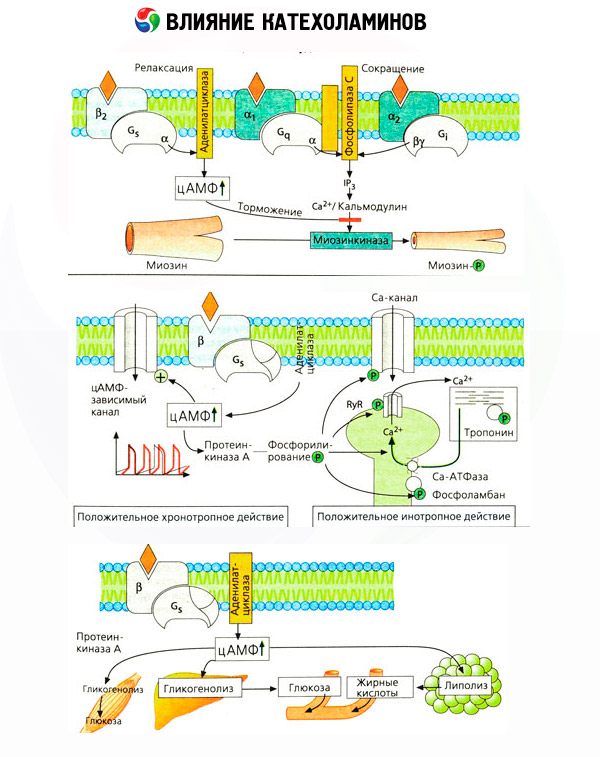
Kombinasi efek langsung dan tidak langsung katekolamin menyebabkan hiperglikemia, tidak hanya terkait dengan peningkatan produksi glukosa hati, tetapi juga dengan penghambatan pemanfaatannya oleh jaringan perifer. Percepatan lipolisis menyebabkan hiperlipidemia dengan peningkatan pengiriman asam lemak ke hati dan intensifikasi produksi badan keton. Penguatan glikolisis di otot menyebabkan peningkatan output dalam laktat darah dan piruvat, yang bersama-sama dengan gliserol dilepaskan dari jaringan adiposa, adalah pendahulu dari glukoneogenesis hepatik.
Pengaturan sekresi katekolamin. Kesamaan produk dan metode respon dari sistem saraf simpatik dan medula adrenal merupakan dasar untuk menggabungkan struktur ini ke dalam tubuh sympathoadrenal sistem pelepasan saraf tunggal dan hormonal link-nya. Berbagai sinyal aferen terkonsentrasi di hipotalamus dan pusat dari sumsum tulang belakang dan medula oblongata dari mana berasal eferen beralih parcel pada badan sel neuron preganglionik terletak di tanduk lateral sumsum tulang belakang pada tingkat VIII serviks - segmen lumbal II-III.
Akson preganglionik dari sel-sel ini meninggalkan sumsum tulang belakang dan membentuk koneksi sinaptik dengan neuron terlokalisasi dalam ganglia dari rantai simpatis, atau sel-sel medula adrenal. Serabut preganglionik ini bersifat kolinergik. Perbedaan mendasar pertama neuron simpatik postganglionik dan medula adrenal kromafin sel terdiri dalam bahwa yang terakhir ditransmisikan ke sinyal yang masuk itu kolinergik neuro-konduksi (postganglionik saraf adrenergik) dan humoral dengan menyorot senyawa adrenergik darah. Perbedaan kedua berkurang saraf postganglionik yang menghasilkan norepinefrin, sedangkan sel-sel medula adrenal - sebaiknya adrenalin. Kedua zat ini memiliki efek berbeda pada jaringan.